


Domain Antigens for Antibody Discovery & Screening
The antibody drug development underwent rapid progress over the past decade, with almost all hot targets being fully covered, making it challenging to identify new targets with good therapeutic potential. Due to the thin margin of new target identification, strategies have changed from “first-in-class” to “best-in-class”, focusing on more delicate, refined drug development work, raising a higher standard for safety and effectiveness. Given that multiple factors, such as function mechanism, epitope, and binding affinity, could affect the clinical performance of antibody drugs, evaluations on these factors at different drug development stages are always required for lead candidate selection. However, this process is often relatively passive, and the success or failure of later screening is highly dependent on whether the initial antibody library(“candidate pool”) contains such excellent candidates. Therefore, it is crucial to obtain a pool of candidates with high specificity and diversity to increase the efficiency of early discovery work.
Domain proteins, or domain antigens, are the essential reagents for establishing early antibody libraries. Whether or not using the appropriate domain antigens during animal immunization will significantly affect the quality of the initial pool and the following antibody screening efficiency. When antibodies targeting specific domains with specific functions or epitope diversity are desired, using domain proteins is a favored option for efficient immunization and screening
Case Study One: Optimizing Mesothelin Antibody Development with Region III Domain
Mesothelin (MSLN) is a critical solid tumor target for multiple cancer indications such as pancreatic cancer, ovarian cancer, and malignant mesothelioma. The drug modality for this target covers monoclonal antibodies, bispecific antibodies, ADCs, etc., with different mechanisms leading to varied development strategies. For example, Eric Hatterer and colleagues found that an antibody (mAb#1) targeting the near-membrane epitope of Mesothelin (anti-Region III) exhibited significantly better ADCC effects than an IgG1 monoclonal antibody [1]. It also showed better ADCP and ADCC effects when combined with a CD47 antibody as a bispecific antibody, compared to another antibody (mAb#2) targeting the far-membrane epitope (anti-Region I) and the benchmark Amatuximab (Figure 1).
|
A. 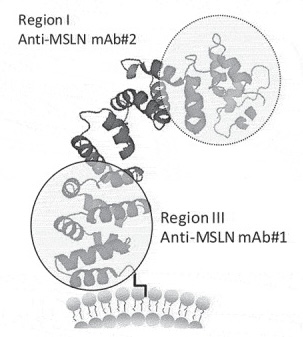 |
B. 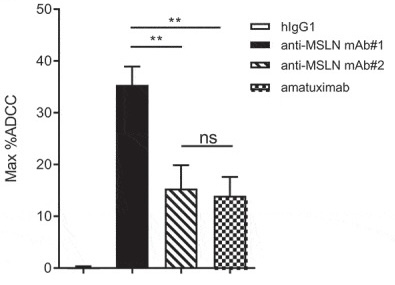 |
|
C. 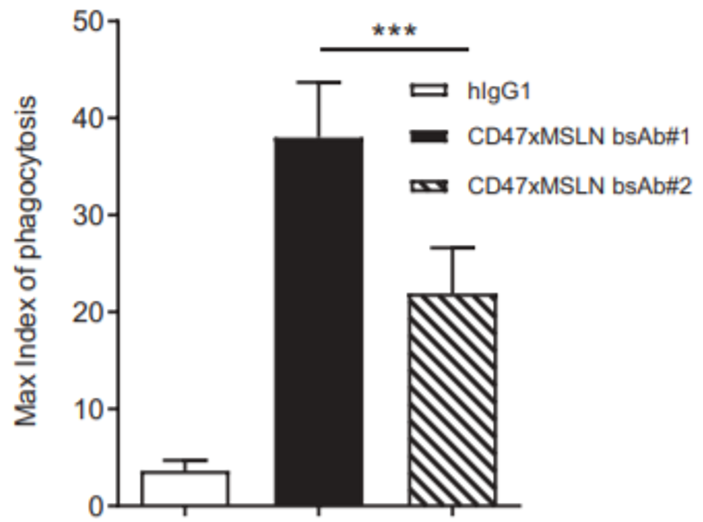 |
D. 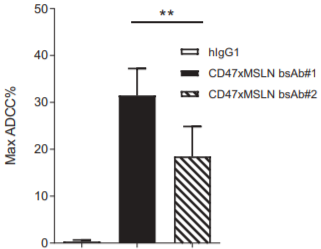 |
Figure 1. Schematic diagram of Mesothelin structure and the impact of different epitopes on antibody in vitro efficacy [1].
When developing Mesothelin antibodies, besides using the full-length extracellular region of MSLN for immunization and screening, considering the use of the Region III domain protein could improve specificity and efficiency, whether alone, or in combination with the full-length extracellular protein for immunization and screening screening. A common concern with domain proteins is ensuring correct conformation when separated from the full length. At KACTUS, we leveraged our extensive experience and knowledge of structural biology to successfully express the Mesothelin Region III domain (#MSL-HM4D1B) with demonstrated good biologic activity (Figure 2).
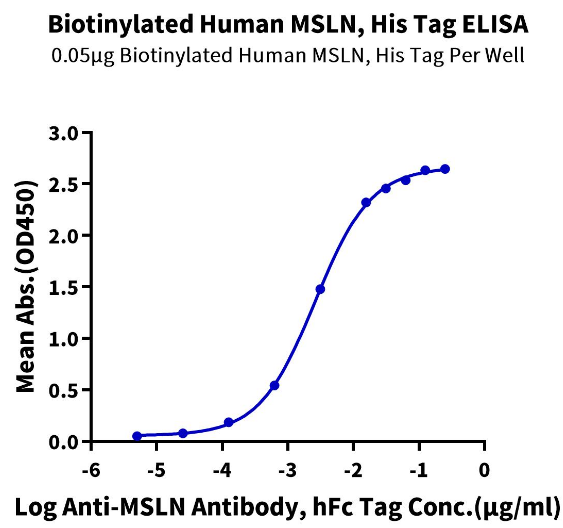
Figure. 2. Immobilized Biotinylated Human MSLN (#MSL-HM4D1B), His Tag at 0.5μg/ml (100μl/well) on the streptavidin precoated plate (5μg/ml). Dose-response curve for Anti-MSLN Antibody, hFc Tag with the EC50 of 2.6ng/ml determined by ELISA.
Case Study Two: Targeting LILRB2 D1D2 Domains for Antibody Development
The tremendous success of anti-PD1/PDL1 antibody drug development has drawn industry attention to evaluating the potential of other immune checkpoints, including the myeloid cell immune checkpoint LILRB2/HLA-G, with several investigational new drugs currently in clinical stages. These antibody drugs usually function by blocking the interaction between LILRB2 and HLA-G, either through steric hindrance or direct competition at the binding epitope.
LILRB2 is composed of four Ig-like C2-type domains. Research by Mitsunori Shiroishi and others has identified that LILRB2 primarily binds HLA-G through its D1 and D2 domains (Figure 3) [2]. This provides a critical insight for antibody development: using the D1&D2 domains of LILRB2 for immunization and screening could enhance the efficiency and specificity of antibodies obtained.
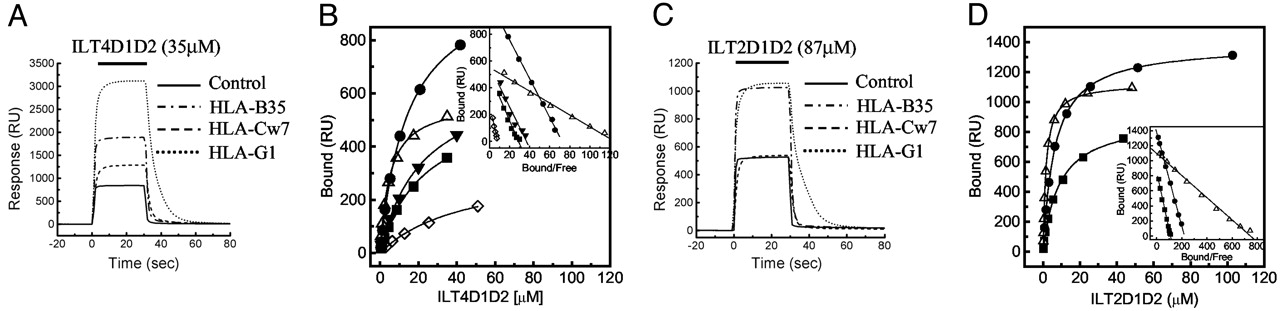
Figure 3. Binding of LILRB1 and LILRB2 with MHC I [2].
Through structural analysis, we believe that the isolated expression of LILRB2 Domain1&2 is highly feasible. Consistent with the feasibility analysis, our LILRB2 Domain1&2 (#LIL-HM12D) demonstrated robust binding activity with the control antibody and was successfully used in blocking experiments (Figure 4).
Immobilized Human LILRB2 Domain1&2, His Tag at 0.5ug/ml (100ul/well) on the plate. Dose response curve for Anti-LILRB2 Antibody, hFc Tag with the EC50 of 16.6ng/ml determined by ELISA (QC Test). |
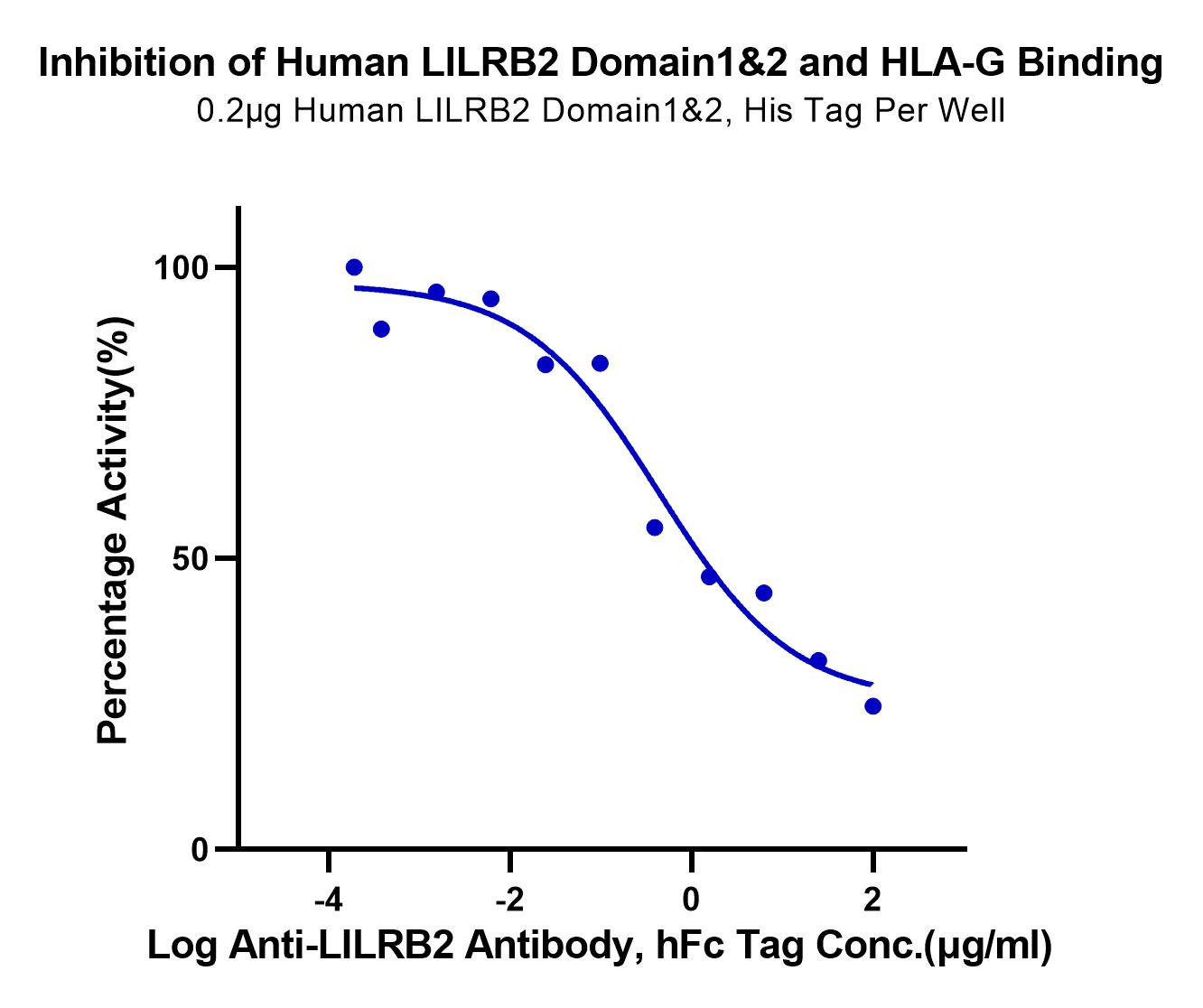 Serial dilutions of Anti-LILRB2 Antibody were added into Human LILRB2 Domain1&2, His Tag: Biotinylated Human HLA-G Complex Tetramer, His Tag binding reactions. The half maximal inhibitory concentration (IC50) is 0.43ug/ml. |
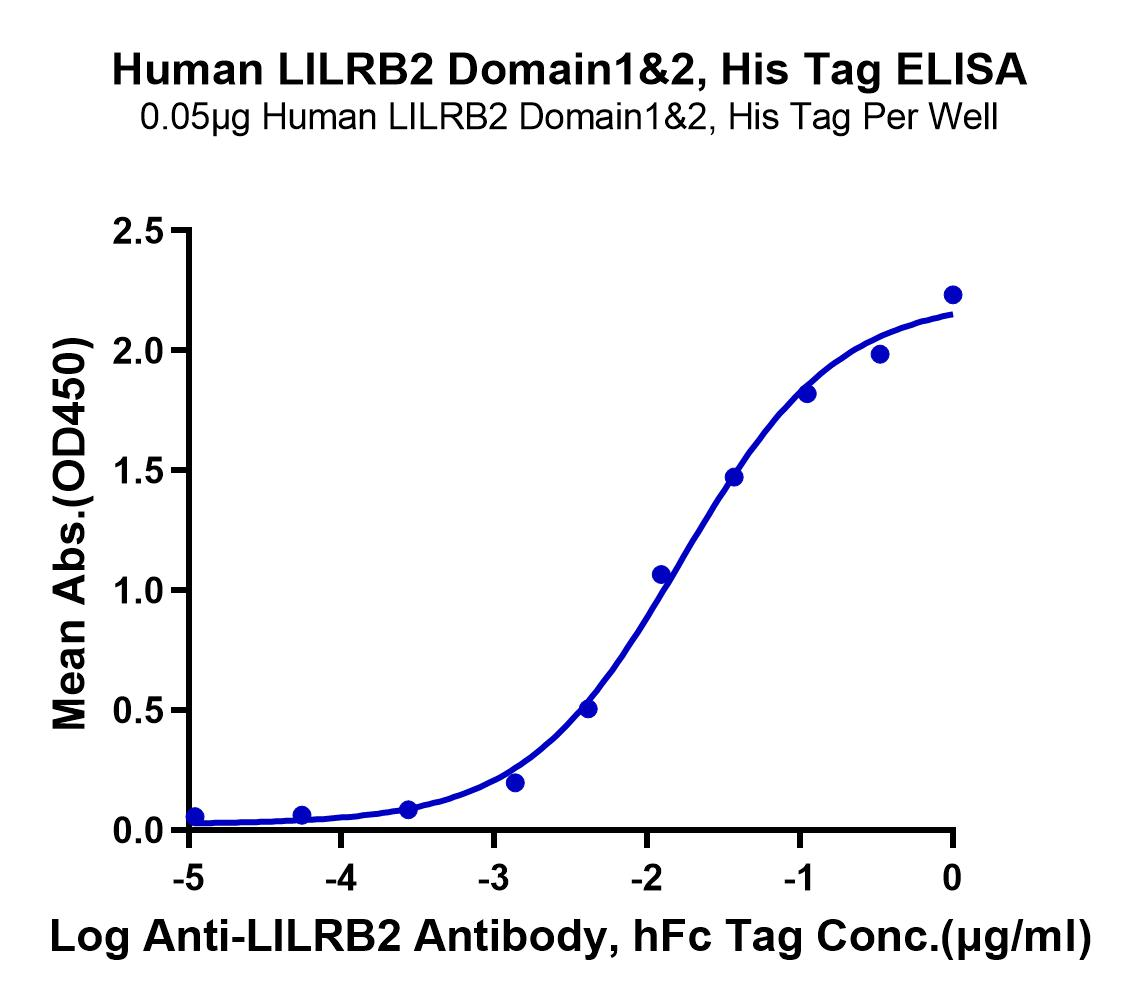
Figure 4. Binding of LILRB2 Domain1 & 2 (#LIL-HM12D) with HLA-G and antibody blocking experiment.
Case Study Three: Overcoming GPC3 Immunogenicity Challenges with VLP-Displayed Domains
Glypican-3 (GPC3) is another promising solid tumor target in recent years, with multiple pipelines under development covering monoclonal antibodies, bispecific antibodies, and ADCs. However, the most significant pain point of GPC3 is the poor immunogenicity, making it challenging for immunization. Current understanding of the GPC3 suggests that high human-mouse homology (up to 95% across the entire ECD) might be a major factor contributing to the low immunogenicity. The high sequence homology avoids producing autoantibodies, a natural characteristic of the immune system as being part of the body's self-protection mechanism. However, this poses challenges for antibody discovery. Sequence comparison reveals that human homology is lower at the C-terminal end of the ECD, making it a potential good epitope for antibody production. Consistent with this, Studies by Mingqian Feng and others demonstrated that antibodies targeting this region, GC33 and YP7, have good therapeutic effects (Figure 5) [3].
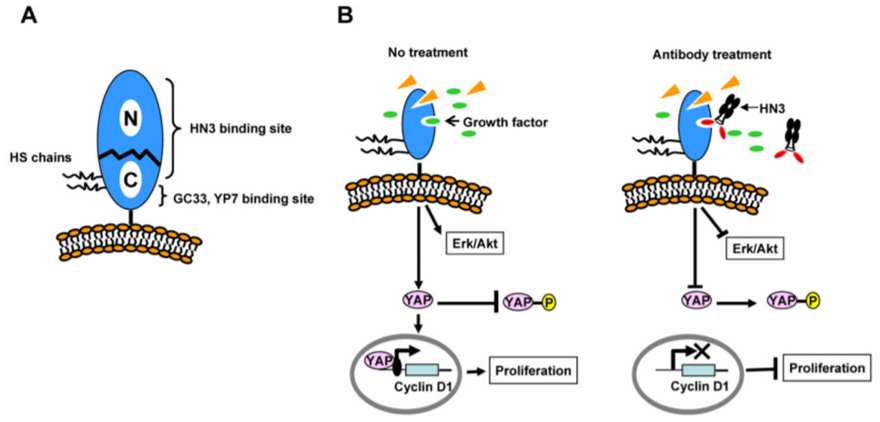
Figure 5. Analysis of GPC3 Antibody Mechanism of Action [3].
Through structure and sequence analysis, we selected Gly510-Asn554 as the target domain and displayed this domain on VLPs to overcome the short length and boost its immunogenicity. Experimental validation showed that the Human GPC3 Protein-VLP (Gly510-Asn554) (#GPC-HE005) binds well to the anti-GPC3 monoclonal antibody and can be used for immunization. Antiserum ELISA testing data showed high titers and excellent binding at a 1:50 dilution with the GPC3 high-expression Huh7 natural cell line, indicating that the VLP-displayed domain closely resembles the natural conformation (Figure 6).
|
A. 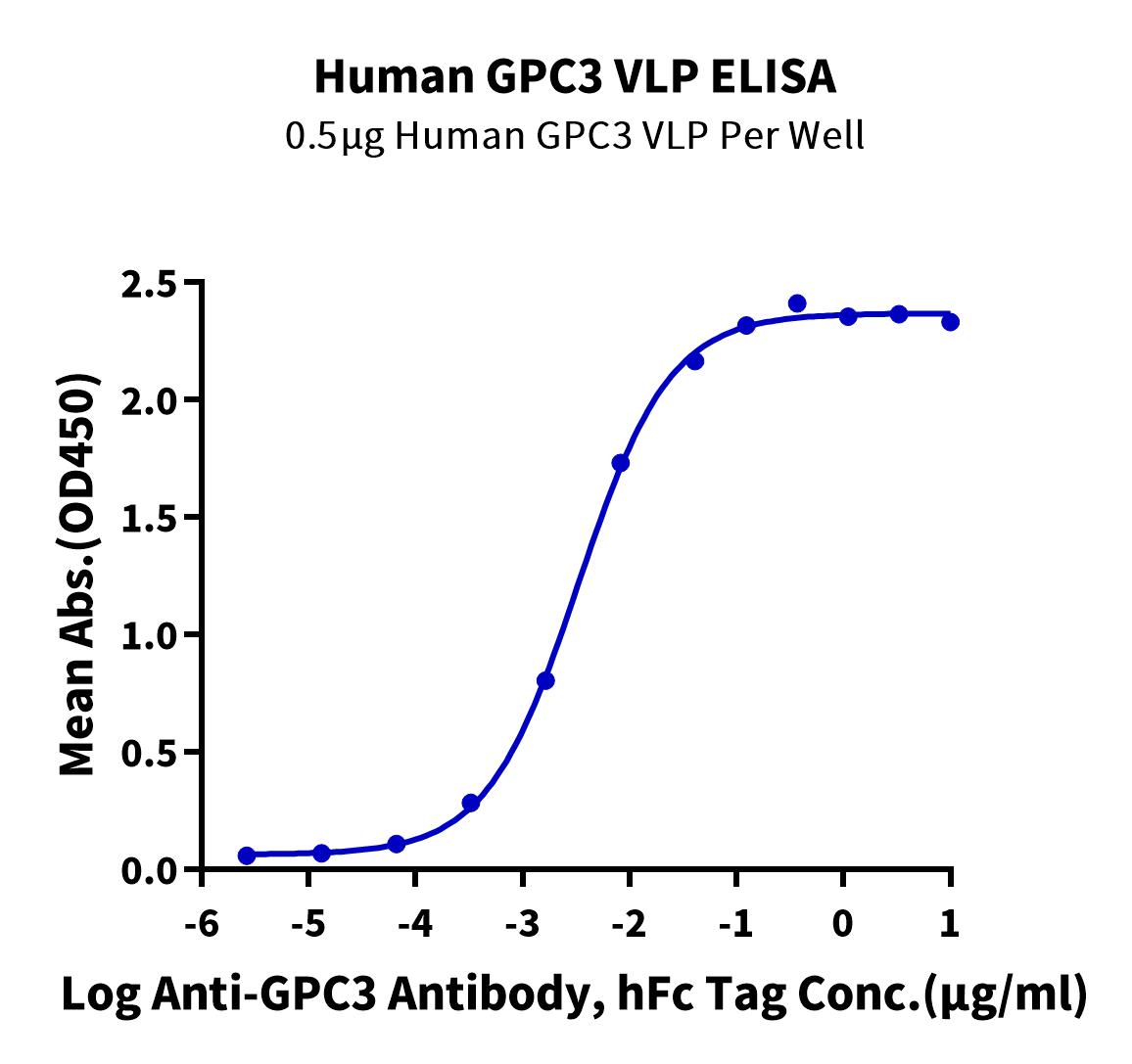 |
 |
C.

Figure 6. (A) Immobilized Human GPC3 VLP at 5ug/ml (100ul/Well) on the plate. Dose-response curve for Anti-GPC3 Antibody, hFc Tag with the EC50 of 3.3ng/ml determined by ELISA. (B, C) Analysis of Immune Antiserum of GPC3 VLP (#GPC-HE005) in ELISA and FACS.
Case Study Four: Enhancing Antibody Specificity with Chimeric pMHCs
Intracellular targets have long been considered to have extensive drug development potential, with TCR therapeutics targeting intracellular targets such as MHC-peptide complexes (pMHC) garnering significant attention. Despite recent advancements, TCR therapeutic studies still face challenges, primarily poor solid tumor infiltration, necessitating further research to improve their efficacy. The success of Kimmatrak, a type of soluble TCR drugs, has successfully overcome the infiltration issue, which has significantly encouraged the field of study and also greatly inspired the development of TCR mimic antibodies, another TCR therapeutic strategy.
The TCR mimic antibody approach aims to utilize the well-established knowledge and technology of antibody research, development, and production platforms in the industry to bypass current technical challenges associated with soluble TCR development. However, this route faces significant challenges in the early antibody discovery phase: pMHCs, the antigen required for TCR-mimic antibody discovery, consist of the HLA α1, α2, and α3 domains, with the antigenic peptide primarily supported by HLA α1 and α2, and effective antibody recognition epitopes concentrated in this area, making it a small, composite epitope. Using the full pMHC for immunization would result in low efficiency in obtaining effective antibodies, as many antibodies produced might target epitopes against HLA α3 and β2M. Direct use of the HLA α1 & α2 & peptide domain strategy is not feasible, as structural analysis shows that, though not directly participate in antigen peptide presentation, HLA α3 and β2M play an important role in maintaining structural integrity, which affects the correct presentation of the antigen peptide between α1 & α2 domains.
To solve this problem, we designed human-mouse chimeric pMHCs, replacing human HLA α3 and β2M with mouse sequences (Figure 7).
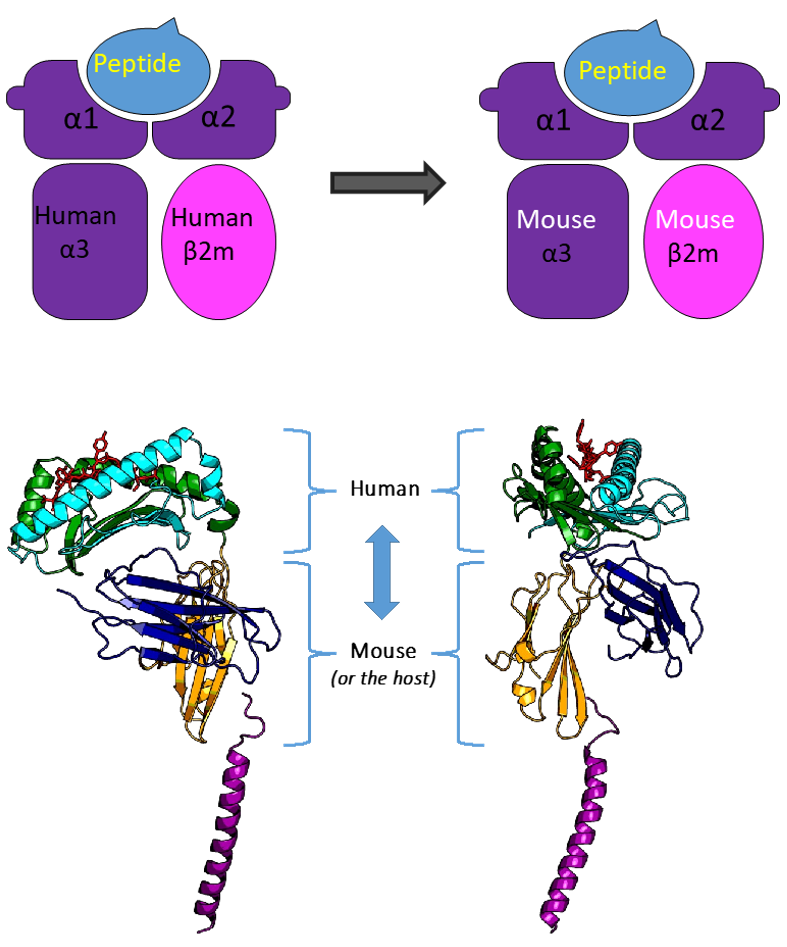
Figure 7. Structure of Human-Mouse Chimeric pMHC.
These chimeric molecules can significantly increase the probability of obtaining antibodies against effective epitopes for mouse immunization. Noticeably, a simple, direct assembly of human-mouse chimeric pMHCs without any structural design or modifications will greatly impair the protein activity (Figure 8, green ELISA marker), suggesting further engineering of the molecules is necessary to maintain the structural and functional integrity of chimeric MHCs. Indeed, by introducing key site mutations, we successfully restored the correct conformation of the chimeric molecule, with ELISA binding assay showing nearly 100% activity for both GP100 and NY-ESO-1 human-mouse chimeric pMHCs (Figure 8).
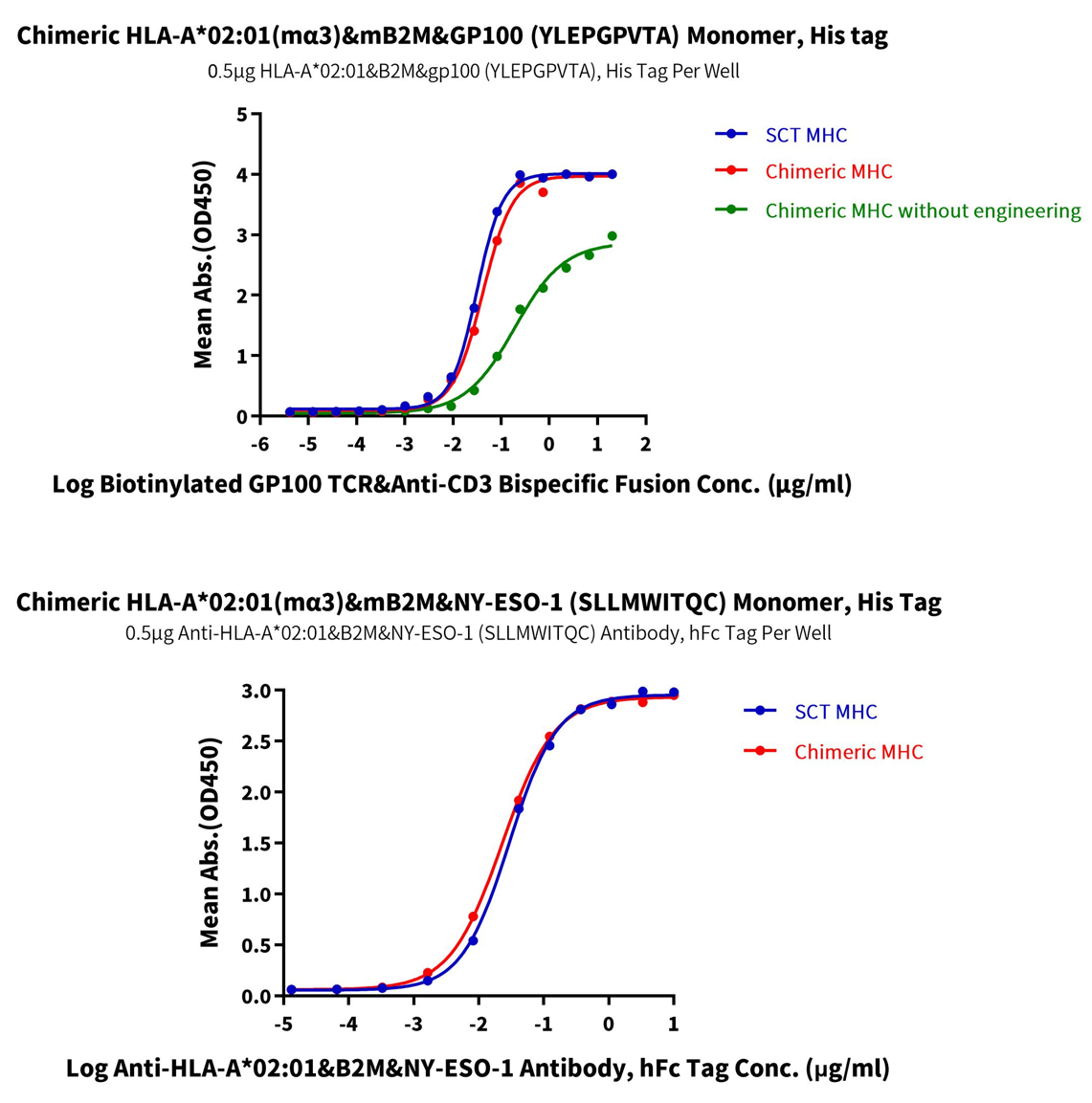
Figure 8. Activity analysis of Human-Mouse Chimeric pMHC.
Conclusion
Despite the remarkable success of antibody drugs, there are still unmet clinical needs, and the competitive market landscape demands higher and more precise requirements for the early development of antibodies. For different targets and mechanisms, it is crucial to adopt differentiated strategies to enhance efficiency and provide ample options for later screening. The application of domain proteins is often a smart strategy in many scenarios, such as precisely targeting functional domains, avoiding the immunodominant effects of ineffective epitopes, precisely targeting near-membrane/far-membrane epitopes, increasing the probability of obtaining cross-reactive antibodies with primates, and enhancing immunogenicity in combination with VLPs. KACTUS is committed to providing high-quality active proteins and aims to optimize antigen proteins for researchers' development goals to enhance development efficiency. If you have any specific antigen requirements for your drug development, please contact us.
References
(1) Eric Hatterer. MAbs. 2020 Jan-Dec;12(1):1739408.
(2) Shiroishi M. Proc Natl Acad Sci U S A. 2003 Jul 22;100(15):8856-61.
(3) Mingqian Feng. FEBS Lett. 2014 January 21; 588(2): 377–382.



